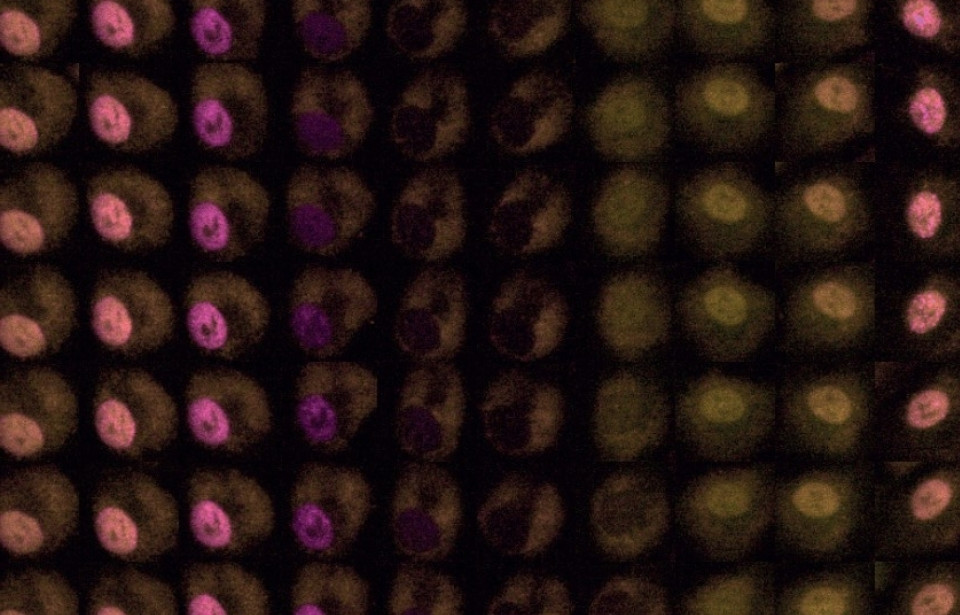
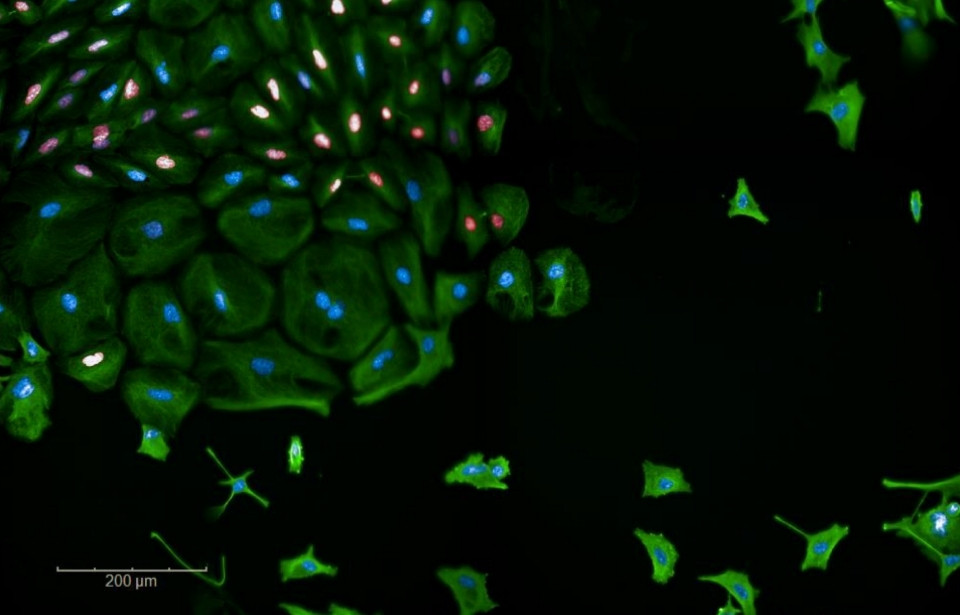
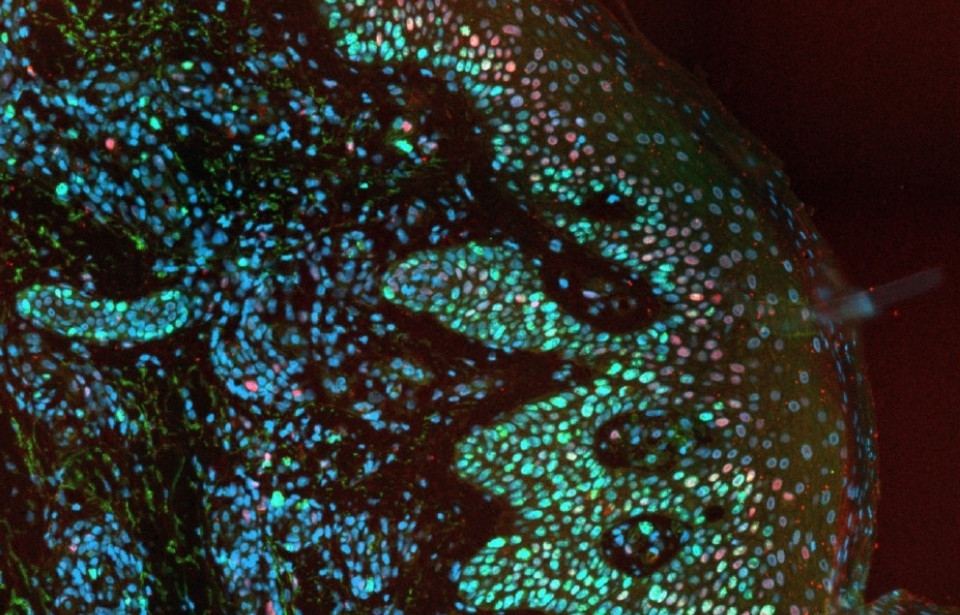
Weston WA, Holt JA, Wiecek AJ, Pilling J, Schiavone LH, Smith DM, Secrier M, & Barr AR. An image-based screen for secreted proteins involved in breast cancer G0 cell cycle arrest (2024). Scientific Data, 11(1). https://doi.org/10.1038/s41597-024-03697-z
Barr AR, Burley A, & Wilkins A. TP53 mutations in urothelial carcinoma: not all one and the same (2024). The Journal of Pathology, 264(2), 125–128. https://doi.org/10.1002/path.6335
Bustraan S, Bennett J, Whilding C, Pennycook BR, Smith D, Barr AR, Read J, Carling D, Pollard A. AMP-activated protein kinase activation suppresses leptin expression indepdently of adipogenesis in primary murine adipocytes (2024). Biochem J. Mar 6;481(5):345-362. Doi: 10.1042/BCJ20240003.
Dragoi CM, Kaur E, Barr AR, Tyson JJ, Novak B. The oscillation of mitotic kinase governs cell cycle latches in mammalian cells (2024). J Cell Sci. Jan 11:jcs.261364. doi: 10.1242/jcs.261364.
Crozier L*, Foy R*, Adib R*, Kar A, Holt JA, Pareri AU, Valverde JM, Rivera R, Weston WA, Wilson R, Regnault C, Whitfield P, Badonyi M, Bennet LG, Vernon EG, Gamble A, Marsh JA, Stable CJ, Saurin A1, Barr AR1, Ly T1. (2023). CDK4/6 inhibitor-mediated cell overgrowth triggers osmotic and replication stress to promote senescence. Molecular Cell. *Joint first; 1Co-corresponding authors.
Wiggins BG, Wang YF, Burke A, Grunberg N, Vlachaki-Walker J, Dore M, Chahrour C, Pennycook BR, Sanchez-Garrido J, Vernia S, Barr AR, Frankel G, Birdsey GM, Randi AM, Schiering C. (2023). Endothelial sensing of AHR ligands regulates intestinal homeostasis. Nature. 2023 Sep;621(7980):821-829. doi: 10.1038/s41586-023-06508-4.
Wiecek AJ, Cutty SJ, Kornai D, Parreno-Centeno M, Gourmet LE, Malagoli Tagliazucchi G, Jacobson DH, Zhang P, Xiong L, Bond G, Barr AR, Secrier M. (2023). Genomic hallmarks and therapeutic implications of G0 cell cycle arrest in cancer. Genome Biology. May 23; 24(1):128. doi: 10.1186/s13059-023-02963-4.
Hughes FA, Barr AR1, Thomas P1. (2023). Patterns of interdivision time correlations reveal hidden cell cycle factors. Elife. Nov 15;11:e80927. doi: 10.7554/eLife.80927. 1Co-corresponding authors.
Swadling JB1, Warnecke T, Morris KL, Barr AR1 (2022). Conserved Cdk inhibitors show unique structural responses to tyrosine phosphorylation. Biophysical J. May 25; S0006-3495(22)00417-9. doi: 10.1016/j.bpj.2022.05.024. 1Co-corresponding authors.
Thomsen I, Kunowska N, de Souza R, Moody AM, Crawford G, Wang YF, Khadayate S, Strid J, Karimi MM, Barr AR, Dillon N, Sabbattini P (2021). RUNX1 controls the dynamics of cell cycle entry of naïve resting B-cells by regulating expression of cell cycle and immunomodulatory genes in response to BCR stimulation. Journal of Immunology. Nov 22;207(12):2976-91. doi: 10.4049/jimmunol.2001367
Pennycook BR, Barr AR1 (2021), Palbociclib-mediated cell cycle arrest can occur in the absence of the CDK inhibitors p21 and p27. Open Biol. Nov 11(11):210125. doi: 10.1098/rsob.210125. 1Corresponding author.
Heldt FS*, Barr AR*, Cooper S, Bakal C, Novak B (2018), A comprehensive model for the proliferation-quiescence decision in response to endogenous DNA damage in human cells. PNAS. Mar 6;115(10):2532-2537. doi: 10.1073/pnas. *Joint first authors
Cooper S, Barr AR, Glen R, Bakal C (2017) Nuclitrack, an integrated nuclei tracking application. Bioinformatics, Jun 20. doi: 10.1093/bioinformatics/btx404.
Asghar US, Barr AR, Cutts R, Beaney M, Babina I, Sampath D, Giltnane J, Arca Lacap J, Crocker L, Young A, Pearson A, Herrera-Abreu MT, Bakal C, Turner NC (2017), Single-cell dynamics determines response to CDK4/6 inhibition in triple negative breast cancer. Clinical Cancer Research, Sep 15;23(18):5561-5572
Barr AR*1, Cooper S*, Heldt FS*, Butera F, Stoy H, Mansfeld J, Novak B, Bakal C1 (2017), DNA damage during S-phase mediates the proliferation-quiescence decision in the subsequent G1 via p21 expression. Nature Communications, Mar 20; doi: 10.1038/ncomms14728 *Joint first; 1Co-corresponding authors.
Barr AR, Heldt FS, Zhang T, Bakal C, Novak B (2016), A dynamical framework for the all-or-none G1/S transition. Cell Systems, Jan 27; 2(1):27-37
Barr AR1, Bakal C (2015), A sensitized RNAi screen reveals a ch-TOG genetic interaction network required for spindle assembly. Sci Rep. Jun 3;5:10564. doi: 10.1038/srep10564. 1Corresponding author.